■ 4.活性連結橋モデル
4.1 活性部位の出現
一つのCa2+イオンがアクチンフィラメント上のトロポニンC(Tn)と結合することによって一つの連結橋(cross-bridge)がアクチンフィラメントと化学的に反応する場が提供される(活性部位active siteの出現)。一つのトロポニンCがこの状態になることをその近傍の一つの連結橋が活性化されたことと同義であると仮定すれば,Ca2+がトロポニンCと結合する化学反応は「活性部位の出現」またはその近傍の「活性連結橋の出現」そのものを表現する化学的過程である。その化学反応は次のように記述される。
Ka
Ca2+ + Tn → TnACT (3-1)
TnはトロポニンC単独の状態を,TnACTはトロポニンCにCa2+が結合した状態を,Kaはこの化学反応の反応速度定数を表わす。Ca2+濃度がTn濃度よりはるかに低値であれば,この化学反応は一次反応速度式で近似される(式3-2)104)。即ち,ある瞬間におけるTnACTの生成速度(d[TnACT]/dt)は定数をKaとしてその時のCa2+濃度([Ca2+])に比例する177)。
d[TnACT]/dt = Ka [Ca2+] (3-2)
等張性収縮においては,ある時点で活性部位近傍にある連結橋だけが回転し力を発生する。それによって心筋が短縮すると今度はその時点での活性部位近傍にある別の連結橋が回転する。このようにして心筋が短縮運動中は活性部位と回転する連結橋の対応関係は時間とともに動的に変化する。一方,等尺性収縮の時には収縮期全体を通じて回転する連結橋の数は増加してゆくが,活性部位と回転する連結橋の対応関係が動的に変化することはない。
心筋の電気的刺激直後に筋小胞体から一気に放出された大量のCa2+が細胞内に拡散し,アクチンフィラメント上のトロポニンCと次々に結合して行く。トロポニンCといったん結合したCa2+は収縮期が終るまで解離せず,収縮期にCa2+は細胞内に取り込まれず,細胞外に排出されず,筋小胞体にも再取り込みされないと仮定すれば,活性部位の濃度は式3-3のようにCa2+放出後の時間の指数関数で増加してゆく。
活性部位の濃度 ∝ 1 – e-Ka・(t – To) (3-3)
収縮期中の任意の時間において活性部位の濃度と細胞内free Ca2+濃度([Ca2+])の合計は一定値であるから,式3-4の様に[Ca2+]はCa2+放出後の時間の指数関数で減少する。
[Ca2+] ∝ e-Ka・(t – To) (3-4)
このようにモデルでは収縮期における細胞内Ca2+濃度の急速な減少は,筋小胞体によるCa2+の汲み上げではなく,Ca2+とトロポニンCの結合反応によるためであると考える。
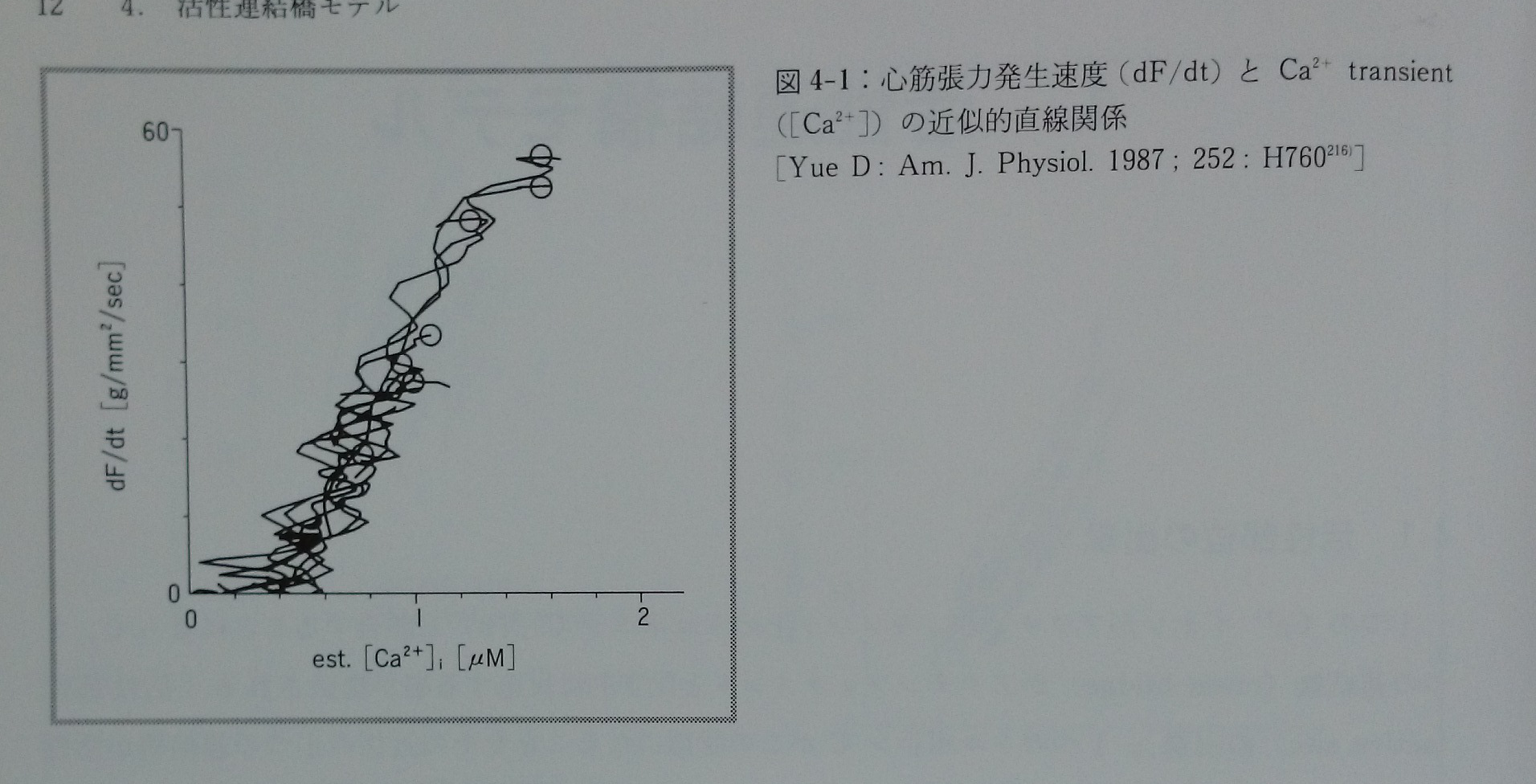
等尺性収縮期の或る瞬間において,Ca2+ transientの値([Ca2+])と心筋の発生張力の一次微分(dF/dt)が直線的関係にある(即ち,dF/dt=k[Ca2+])ことをYue216)が報告した(図4-1)。この知見はモデルから容易に説明できる。即ち,心筋発生張力の時間微分(dF/dt)は活性部位濃度の時間微分(d[TnACT]/dt)に比例することから,式3-2は式3-5となる。
dF/dt ∝ Ka [Ca2+] (3-5)
ある瞬間における心筋発生張力の一次微分とその時の[Ca2+]が直線的関係となることがモデルから理論的に説明される。比例定数はKaと密接に関係する。従って,モデルで仮定したCa2+とトロポニンCの結合反応の一次反応速度式による近似は妥当である。
4.2 心筋発生張力の基本方程式
式3-2から収縮期心筋の力ー長さー時間関係(myocardial force-length-time relationship)を表す式3-6を導く168)。
F = E(t)・(L – Lo) (3-6)
E(t) = (Ec/α)・{1 – e-Ka・(t – To)} (3-6′)
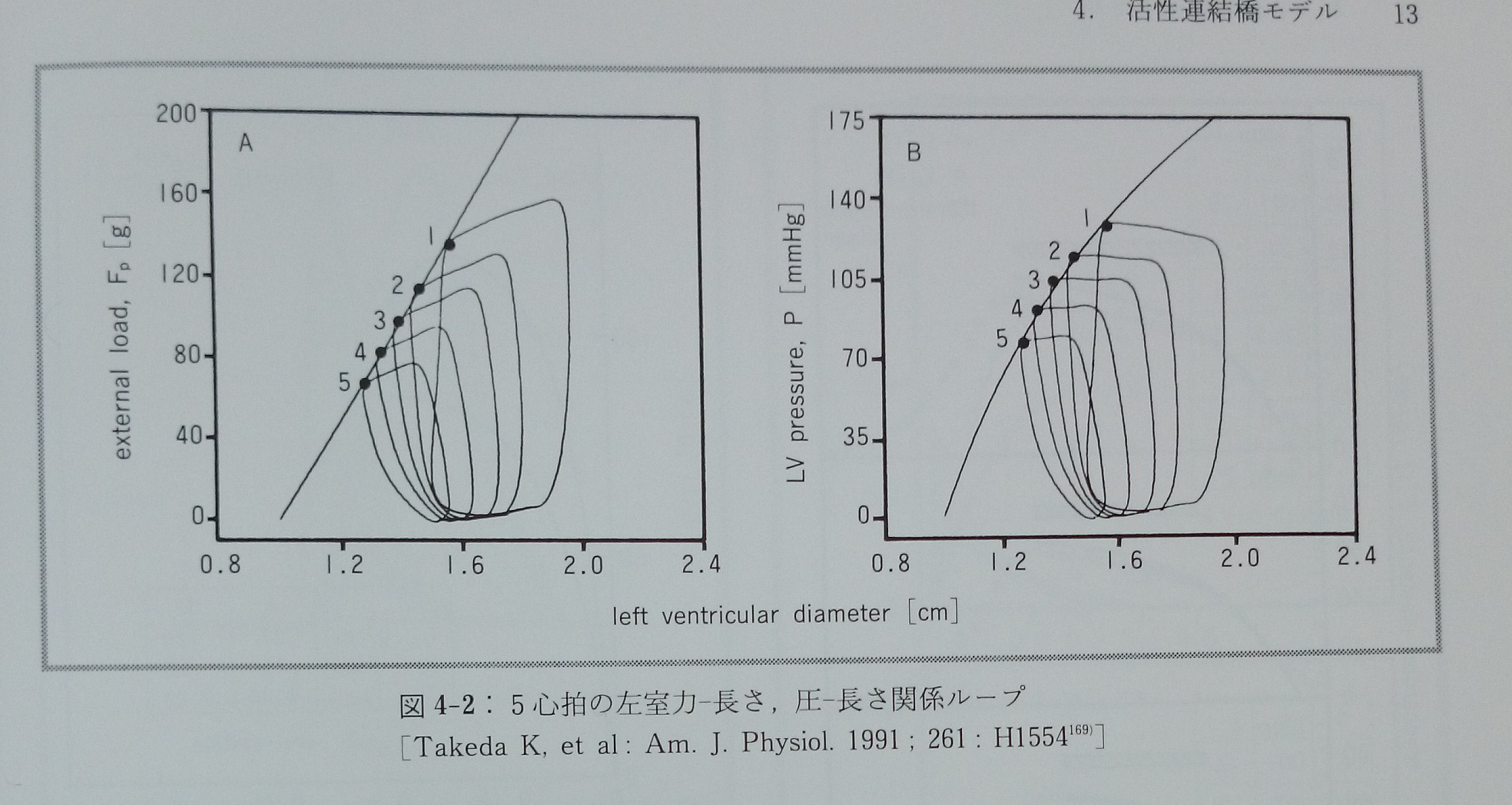
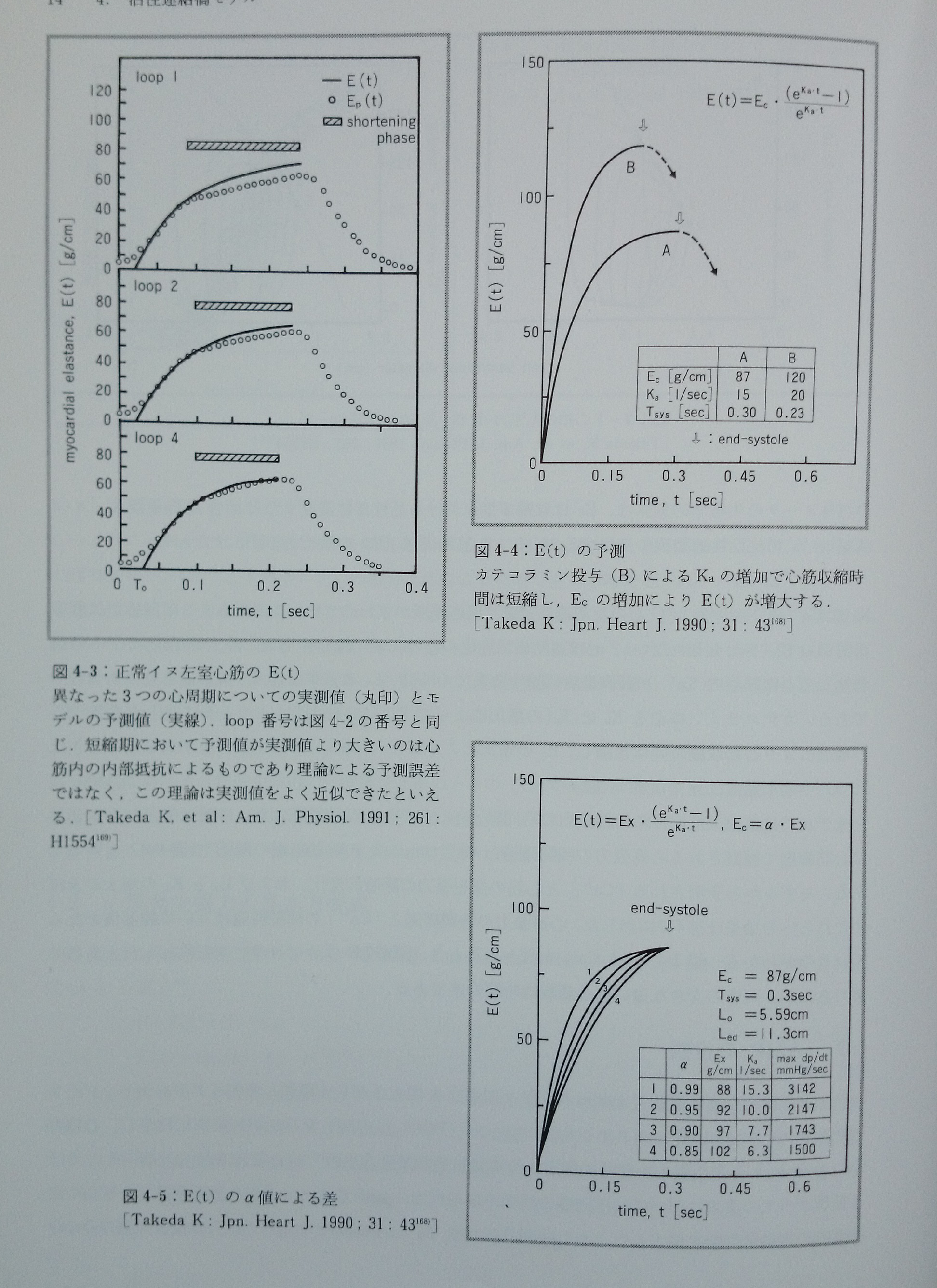
式3-6と3-6’が活性連結橋モデルにおける収縮期心筋力学の基本方程式である。ここでは心筋収縮力(F)は二つの独立変数(心筋長Lと時間t)の関数であり,E(t)は心筋細胞内の活性部位濃度または活性連結橋濃度に対応し,時変心筋弾性(time-varying myocardial elastance)と表現し得る。正常イヌ左室心筋の異なった心拍(図4-2)のE(t)を図4-3に示す169)。
心筋収縮力Fは少なくとも5つのパラメ-タ-(Ec, Ka, α, To, Lo)によって修飾される。これらのパラメ-タ-の生物学的意味は,Ecは収縮末期における活性部位濃度または活性連結橋濃度[α・σ(Ca2+T)],単位活性連結橋の発生する力(f),心筋断面積(S)の積であり169),即ち,
Ec = α・σ(Ca2+T)・f・S (3-7)
KaはCa2+とトロポニンCとの反応速度定数(反応速度が極めて早いことから,この反応には酵素が関係しているかも知れない),αは連結橋活性化の際のCa2+利用率(Ca2+のトロポニンCへの親和性),Toは細胞内Ca2+が最高濃度に達するまでの時間,Loは基準心筋長(無機能心筋長を含む)である。catecholamineによるKaとEcの増加によってE(t)の形は図4-4の如く変化する168)。Kaの増加により心筋収縮時間は短縮し,Ecの増加によりE(t)が増大する。またα値が低下した時はE(t)の増加速度は遅くなる168)(図4-5)。
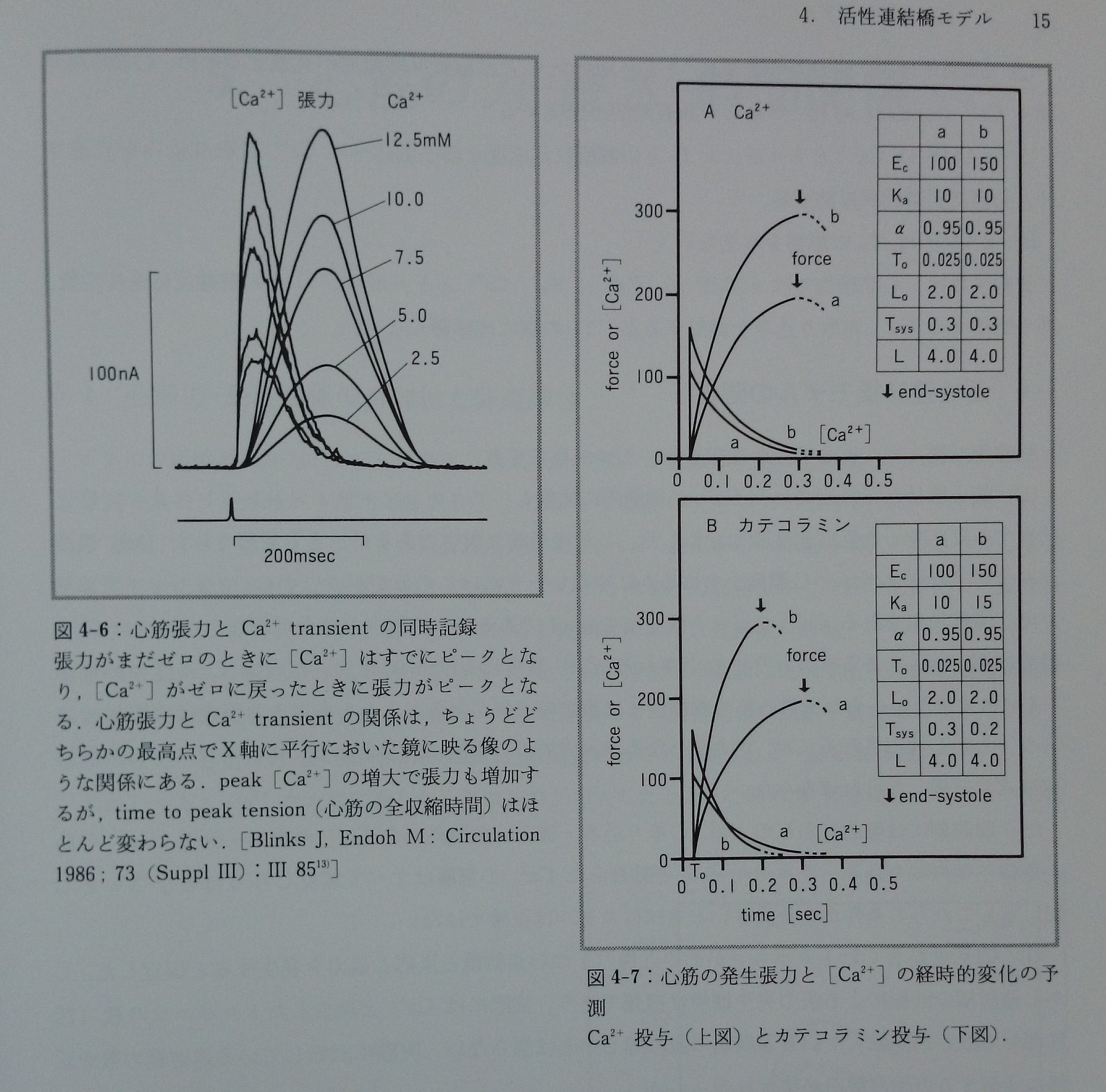
モデルでは収縮期における心筋張力の時間経過と[Ca2+]の時間経過は互いに鏡面像となる。これは心筋細胞で観察される心筋張力の時間経過とCa2+ transientの時間経過の関係13)(図4-6)と同じである。モデルから予測される[Ca2+]と心筋の発生張力の経時的変化,およびEcとKaの増大が及ぼすこれらへの効果は図4-7に示した。心筋張力の時間経過と[Ca2+]の時間経過は互いに鏡面像となっているのが分かる。図4-7AはCa2+を投与した時,図4-7Bはcatecholamineを投与した時の効果であるが,両者の大きな違いは心筋収縮時間の差である。
4.3 cAMPの役割
ラットの摘出心室乳頭筋でcyclic AMP (cAMP)を増大させる4種類の薬剤(adrenaline, isoproterenol, cAMP, phosphodiesterase inhibitor (PDE阻害剤))をそれぞれ個別に投与し,心筋のCa2+ transientと発生張力を同時に測定した実験結果が栗原と小西99)から報告されている。そこでは4種類全ての薬剤においてほぼ同様の結果が得られた。peak Ca2+ transientと発生張力はともに投与薬剤の容量依存性に増大した。また時間分析ではCa2+ transientのピークからの減衰時間(decay time to light)は薬剤投与で平均30.8%程度有意に減少し,心筋収縮時間(time to peak tension)は平均15.0%減少し,心筋拡張時間(relaxation time)は平均35.5%有意に減少した。しかしCa2+ transientのピークまでの時間(time to peak light)の減少は平均8.3%と変化が少なかった。peak Ca2+ transientは筋小胞体から放出された[Ca2+]であり,Ca2+ transientのピークからの減衰時間と心筋収縮時間はCa2+とトロポニンCとの結合反応速度定数(Ka)によって主に決定され,心筋拡張時間はCa2+とトロポニンCとの解離反応速度定数によって主に決定され,Ca2+ transientのピークまでの時間はToである。
従って,以上の実験結果はこのモデルでは次のように解釈される。1)cAMPは筋小胞体から放出される[Ca2+]を容量依存性に増加させ,心筋発生張力がcAMPの容量依存性に増大する。2)cAMPはKaを増大させCa2+とトロポニンCとの結合反応速度が亢進するため,心筋収縮時間やCa2+ transientのピークからの減衰時間が短縮する。3)cAMPはCa2+とトロポニンCとの解離反応速度と筋小胞体へのCa2+再取り込みを亢進させ,心筋拡張時間が短縮する。4)cAMPはToに影響を与えない。要約すれば,心筋細胞ではcAMPが[Ca2+],Ka,Ca2+とトロポニンCとの解離反応速度定数,筋小胞体へのCa2+再取り込みを調節するがToの値には影響しな
い。
4.4 活性連結橋モデルの限界
活性連結橋モデルにも明らかな限界が幾つか存在する。1)筋小胞体から放出されたCa2+が細胞内に拡散し,アクチンフィラメント上のトロポニンCと結合する反応を一次反応速度論で近似した。これは単純な仮定であるが,もし放出されたCa2+濃度が結合可能なトロポニンC濃度よりはるかに少ない時にはこの仮定が成立する。フェレット乳頭筋では通常放出されるCa2+のピーク濃度は1 μmolであるが,細胞内の結合可能なトロポニンCの濃度は70 μmolであるとされ100),放出されるCa2+濃度は結合可能なトロポニンC濃度の僅か1.4 %にすぎない。また正常心筋の収縮性には相当の予備があることからも以上の仮定は実際的であろう。しかし放出されるCa2+が大量となる状況ではこの仮説は無効となる。この時には二次反応速度式で近似しなければならない。2)収縮期には細胞外からのCa2+の取り込み・流入,細胞外へのCa2+の放出・流出,Ca2+の筋小胞体への再取り込み,トロポニンCと結合したCa2+の解離は全て無視し得るほど小さいとした。もしこれらの条件が十分大きい時はにモデルは正確ではない。3)Ca2+と結合したトロポニンがその近傍の一つの連結橋と反応し張力を発生すると仮定した。しかし連結橋の回転による張力発生機構は複雑であり,実際にはCa2+と結合したトロポニンの数(活性部位の数)が回転している連結橋の数に等しいとは限らない。実際に回転している連結橋の数を正確に予測する方法の解明が待たれる。4)筋小胞体から放出されたCa2+が細胞内に拡散しトロポニンと結合するが,この反応速度定数は物理的なCa2+の拡散能によって単純に決定されるかどうかは不明である。この結合反応速度は非常に早く,少なくともcatecholamine,cAMP,PDE阻害薬,甲状腺機能異常,体温によっても影響を受ける。単純な細胞内拡散に加えてこの結合反応には何らかの酵素が関与しているかも知れない。この結合反応速度を支配する詳細な機構の解明が待たれる。